Uma micorriza (do grego myco , “fungo” e riza , “raiz”, termo introduzido em 1885 pelo botânico Albert Bernhard Frank ) é o resultado da associação simbiótica , chamada de micorrização , entre fungos e raízes de plantas . As micorrizas são um dos principais componentes do edafão e da rizosfera .
Neste associação não específica, geralmente, os esporos de fungos micorrízicos ou micorrízicos fungo (do grego myco , rhiza e genos "para engendrar", literalmente, o que dá origem a uma micorrizas) são disseminados pelo vento ( anemochoria ), pela chuva ( hidrocoria ), ou por excrementos de animais ( endozoocoria ), germinam e dão origem às hifas do micélio que colonizam as raízes de uma planta. Essas hifas radiculares são distintas das hifas extrarradiculares, que também crescem fora da raiz por vários centímetros, explorando o solo ao redor do sistema radicular da planta hospedeira. O que é comumente chamado de fungo, que é colhido com o pé e o chapéu, é apenas a "frutificação" do micélio, o esporóforo , onde ocorre a reprodução sexuada . As hifas aparecem como filamentos finos, capazes de explorar um volume muito grande de solo (mil metros de filamentos miceliais por um metro de raiz).
A relação micorrízica é do tipo simbiótica , mas um desequilíbrio na relação pode ser induzido por uma fraqueza de um dos dois parceiros, podendo então a associação deslizar ao longo do continuum mutualismo-parasitismo. O fungo também pode ajudar a reciclar a necromassa de seu hospedeiro, para o benefício de seus dois descendentes.
Enquanto a superfície de exploração usada pela planta é multiplicada por 10 pelos cabelos ativos da raiz apenas durante o período de germinação , ela é multiplicada por 10.000 graças às micorrizas (filamentos com diâmetro de um centésimo de milímetro) que têm um custo energético de pega até cem vezes menos do que as raízes. Os cabelos da raiz permanecem funcionais em alguns grupos de plantas adultas não micorrízicas (menos de 10%) que perderam secundariamente a associação fúngica, mais de 90% sendo colonizados por essas hifas do fungo simbionte que fornecem a maior parte da absorção de água e nutrientes, e aumentar dez vezes o crescimento da planta. As estimativas sugerem que existem cerca de 50.000 espécies de fungos que formam associações micorrízicas com 250.000 espécies de plantas, 80% dessas associações sendo endomicorrizas arbusculares. Parece que as plantas não micorrízicas de ambientes pioneiros (ausência de fungos), úmidos e / ou ricos, nos quais o suprimento hidromineral não necessita de micorrizas, "precisaram compensar a ausência do fungo auxiliar, desenvolvendo estruturas radiculares que mimetizam micélio. filamentos e sua alta eficiência na colonização de grande volume de solo ”.
Pesquisas atuais mostram uma associação mais ampla, ao nível das micorrizas, com o conceito de microbioma micorrízico, denominado rizomicrobioma, que também envolve bactérias.
O simbionte original (um glomeromiceto ao que parece) teria surgido há cerca de 500 milhões de anos no Paleozóico , provavelmente ao mesmo tempo que as primeiras plantas terrestres. Os fósseis da flora de Rhynie (por exemplo, Aglaophyton (in) , Rhynia (in) ), com cerca de 400 milhões de anos, contêm glomales morfologicamente idênticos às micorrizas , associadas aos rizomas . Isso sugere que as micorrizas foram o instrumento de uma colonização acelerada das terras emergidas, por sua capacidade de extrair água e minerais do solo.
Essas primeiras associações também foram capazes de permitir a constituição de um solo mais resistente às intempéries, com melhor armazenamento de água, melhorando a resistência das plantas ao estresse hídrico e ao frio ou à falta de nitrogênio (no abeto por exemplo), sua resistência ao vento e o escoamento, bem como talvez a sua resistência a excessivas intensidades luminosas, como é o caso dos micoficobiontes de certas algas da costa (estas simbioses, principalmente com os ascomicetes , parecem ter surgido secundariamente).
Associações micorrízicas (arbuscular e ectomicorrízica) também existem em ecossistemas frios (temperatura média abaixo de 15 ° C), onde são ativas todo ou parte do ano (dependendo da temperatura do solo), envolvidas em certas adaptações de resistência ao gel e favorecem a aquisição de nutrientes a baixa temperatura em fungos micorrízicos. Sem dúvida, contribuem para uma melhor sobrevivência do fungo em solos congelados no inverno, onde os micro-fungos sofrem esse fator de seleção adicional. Essas simbioses frias também beneficiam gramíneas como a cevada ( Hordeum ), o que melhora seu acesso ao fósforo do solo .
Estes simbioses corresponder a uma “ macro salto evolutivo ” (salto evolutivo) uma vez que as funções de um parceiro estão associadas com as funções do outro, com efeitos funcionais multiplicado. Na verdade, passamos do estado de alga e fungo para o de planta micorrízica terrestre (o que vai contra o gradualismo darwiniano ).
Atualmente, 85% dos Archegoniates , além dos Hepáticos , são endomicorrízicos por Glomales . Isso supõe que a simbiose com os Glomales é a mais antiga entre os Archegoniates e que teria permitido a irradiação impressionante destes últimos (diversidade, lignificação ...). 80% das plantas vasculares são colonizadas por fungos endomicorrízicos, enquanto as ectomicorrizas são frequentemente associadas a árvores florestais lenhosas.
As outras famílias de Glomeromycetes ( Acaulosporaceae e Gigaspora ) apareceram mais tarde por volta de -250 / -230 milhões de anos atrás. Eles têm capacidades superiores para a exploração dos recursos minerais do solo.
As ectomicorrizas , por sua vez, podem ter surgido no Cretáceo , mas os fósseis mais antigos conhecidos datam apenas do Eoceno . Eles permitiram a colonização de solos antes desfavoráveis. Foi também nessa época que surgiram as simbioses de fixação de nitrogênio . A simbiose ectomicorrízica apareceu muitas vezes de forma independente em diferentes clados de fungos (80 vezes em glomeromicetos , ascomicetos e basidiomicetos ) e espermafitos (12 vezes em gimnospermas e angiospermas ). Essa frequência mostra o sucesso evolutivo dessas associações dos ancestrais dos fungos ectomicorrízicos que viviam de matéria orgânica morta no solo em um modo de vida saprofítico . O estudo comparativo dos genomas dos fungos ectomicorrízicos e saprofíticos atuais mostra que as células micorrízicas perderam muitas exoenzimas que garantem a nutrição carbonácea autônoma dos saprófitos, porque são nutridos com açúcares pela planta hospedeira, mas mantiveram as enzimas (peroxidases, lacases, fenol-oxidases fúngicas, como tirosinases) que exploram os recursos de nitrogênio e fosfato de resíduos orgânicos (lignina, compostos fenólicos, celulose) necessários para o bom funcionamento da planta.
Algumas plantas com simbioses micorrízicas mais recentes (do ponto de vista evolutivo), também podem contrair uma associação com micorrizas arbusculares. Portanto, parece que houve inovações evolutivas neste tipo de simbiose. Essas inovações, sem dúvida, explicam o número limitado de plantas capazes de contratá-los.
O aparecimento de ectomicorrizas foi correlacionado duas vezes com a radiação evolutiva das plantas:
Mais recentemente Ainda outras formas de endomicorrizas e ectomicorrizas de pelotão surgiram, especialmente em Ericales , com o fenômeno da micoheterotrofia : seus simbiontes fúngicos têm capacidades saprofíticas ainda mais fortes , permitindo à planta re-ingerir carbono orgânico diretamente através do fungo, em solos onde a atividade de mineralização é muito baixa ( pântanos , altas montanhas, ecossistemas frios, etc.): a simbiose permite um acoplamento de níveis tróficos .
Vários grupos de plantas (menos de 10%) não são micorrízicos. Eles perderam secundariamente a associação e ilustram o fenômeno da neotenia . São essencialmente plantas de ambientes pioneiros (ausência de fungos) ou ricos. Por exemplo, espumas adaptadas a ambientes hostis, onde podem secar e onde um fungo sobreviveria mal. Eles adquiriram estruturas anatômicas que lhes permitem resistir à dessecação , mas, correlativamente, seu tamanho permanece pequeno. Também são angiospermas ( Brassicaceae , Chenopodiaceae , Polygonaceae , Proteaceae , Caricaceae, etc.) que se adaptaram com mais frequência a ambientes ricos, nos quais o suprimento hidromineral não requer micorrizas.
As micorrizas estão na origem dos ecossistemas mais complexos, em particular nas florestas e, em particular, nas florestas tropicais que muitas vezes vivem e evoluem em solos ingratos e por vezes inférteis. Seus micélios formam redes miceliais interconectadas que influenciam o funcionamento dos ecossistemas (ciclos biogeoquímicos, composição de comunidades de plantas, nutrição carbonácea de mudas durante seu desenvolvimento, modificação da competição, etc.), permitindo ou aumentando fluxos significativos de carbono orgânico e minerais ( nitrogênio , fósforo , água ...) através do solo (em média 30 a 40% dos minerais capturados pelas margens da rede micelial são retrocedidos para a raiz, esta última fornecendo 20 a 40% dos carboidratos fotossintetizados para o fungo). Eles constituem um dos elementos mais dinâmicos da simbiose micorrízica e desempenham um papel essencial no funcionamento e estruturação das comunidades de plantas. Essas transferências são tão eficazes que desafiam o conceito de especiação por competição por nutrientes entre as plantas de um ecossistema, em particular para a captura de fosfatos pelas raízes (eles tornam possível prescindir de fertilizantes fosfatados), para resistência ao cálcio tóxico (algumas plantas calcicolas toleram o cálcio graças às hifas miceliais rejeitando ativamente os íons Ca 2+ para fora ou imobilizando-os na forma de cristais de oxalato de cálcio que precipitam em vacúolos , idioblastos ou nas paredes das hifas) ou para resistência à seca . No entanto, ainda são pouco explorados na horticultura , agricultura e silvicultura , ou para a descontaminação de certos solos poluídos.
Certos grupos de fungos são provavelmente espécies-chave ou mesmo “espécies engenheiras” que influenciam os principais processos ecológicos do solo. Eles são considerados pelos cientistas do solo como elementos essenciais da diversidade das comunidades, o que é um fator de estabilidade e equilíbrio ecológico. Muitos grupos-chave encontrados no solo (bactérias e fungos micorrízicos em particular) podem se conectar às plantas (pelo menos 90% das famílias de plantas terrestres estão envolvidas) por meio de associações micorrízicas arbusculares e desempenham sinergias essenciais para a sobrevivência e a produtividade das plantas, ajudando a formar uma rede ecológica essencialmente subterrânea, que alguns biólogos chamam de wood-wide web (em referência à “ World wide web ”). Em 1997, a pesquisadora canadense Suzanne Simard foi a primeira a demonstrar essa rede com a transferência micorrízica de carbono entre árvores em condições naturais.
A maioria dos fungos micorrízicos são suspeitos de ter vários hospedeiros ou mesmo uma ampla gama de hospedeiros (cada planta sendo comumente associada a várias dezenas de fungos micorrízicos diferentes), o que parece ser confirmado em ambientes naturais , mas estudos feitos sobre eles Solos aráveis cultivados mostram, no entanto, que a diversidade de fungos micorrízicos é "extremamente baixa em comparação com solos florestais" .
A colonização do sistema radicular, o potencial “micorrizogênico” do solo e a “dependência micorrízica” das plantas estão inversamente correlacionados com o conteúdo da solução do solo em íons fosfato ; além disso, esse resultado não está relacionado a nenhuma forma de fertilizante fosfatado, seja orgânico ou inorgânico, uma vez que as plantas só absorvem íons em solução. O enriquecimento desta solução torna-se diretamente responsável pelo fato de a planta bem nutrida não promover mais o desenvolvimento de micorrizas. Em certas situações, os níveis de fósforo alcançados tornam-se incompatíveis com a instalação de micorrizas.
As micorrizas também interagem com várias bactérias do solo (incluindo Pseudomonas ) que podem ser patogênicas (as micorrizas protegem a planta contra seus patógenos, por exemplo, emitindo antibióticos), mas que também são chamadas de "bactérias auxiliares da micorrização" (em inglês MHB: Mycorrhizal Helper Bacteria ), pois desempenham um papel tão importante.
As micorrizas também interagem com outras micorrizas e outros fungos e com certos predadores aéreos e parasitas de plantas:
Numerosos experimentos de micorrização controlada mostraram que a regeneração natural ou artificial se beneficia da presença ou inoculação de fungos simbiontes adaptados às plantas e ao contexto. Por outro lado, a cultura de trufas só poderia ser exportada para a Nova Zelândia com plantas hospedeiras micorrízicas. Da mesma forma, as árvores de Natal europeias só cresciam lá com seus simbiontes. Alguns simbiontes são espetacularmente eficientes: é um aumento de 60% no volume total de Pseudotsuga menziesii que foi permitido em 10 anos no florestamento de plantas ectomicorrízicas em viveiros com uma cepa Laccaria bicolor conhecida como S238N, em comparação com árvores não inoculadas.
Precauções : Existe o risco de competição com espécies locais, ou mesmo poluição genética . Vários acompanhamentos mostraram que as plantas inoculadas em viveiros geralmente perderam esse simbionte para outras do solo local, mas pode nem sempre ser o caso.
Ao contrário da crença popular, a relação micorrízica não é exclusivamente simbiótica porque as transferências de substâncias do fungo para a planta nem sempre aumentam o valor seletivo (que é o critério para definir a simbiose ). Os benefícios, que variam muito dependendo do genótipo dos parceiros e do ambiente, são difíceis de avaliar, especialmente para plantas perenes. Eles podem se beneficiar da simbiose apenas por curtos períodos em diferentes momentos de suas vidas. Na verdade, existe um continuum de relações entre os parceiros, da simbiose ao parasitismo, passando pela saprotrofia (a natureza da relação pode ser descrita em função da relação custo / benefício).
Por exemplo, Tricholoma matsutake é simbiótico, parasita ou saprofítico, dependendo de seu estágio de desenvolvimento e das condições ambientais; orquídeas micorrizas são parasitas de plantas lenhosas e ninhos de pássaros neotti são parasitas de suas micorrizas.
Na verdade, a micorrização é continuamente instável devido aos conflitos de interesse entre os parceiros e à pressão de seleção que empurra cada parceiro para ser o mais parasita possível, o que explica as muitas interações possíveis: trocas de nutrientes (a planta fornece açúcares e lipídios , micélios fúngicos fornecem água e sais minerais que extraem do solo, às vezes até vários metros da raiz), de fatores de crescimento ( hormônios , vitaminas ), os alomônios fornecem proteção. Muito parasitismo pode levar ao colapso da associação. Com o tempo, a "simbiose" teria surgido e se quebrado várias vezes: a associação é reversível. Os fungos saprofíticos podem, portanto, ser simbiontes que perderam seu hospedeiro. Assim, em certas associações micorrízicas, um dos parceiros parece explorar o outro (seja por natureza ou dependendo das condições). O termo “explorador” é aqui entendido como um indivíduo que obtém um benefício (aumento do valor seletivo) sem reciprocidade. Podemos citar como exemplo as plantas micoheterotróficas (mais de 200 espécies conhecidas em orchidaceae , gentianaceae , burmanniaceae, etc.), plantas com capacidade fotossintética zero ou reduzida que obtêm seu carbono de seu parceiro micorrízico, ele mesmo obtendo-o de 'um parceiro de clorofila através da rede micelial.
A exploração de um dos sócios da simbiose pelo outro supõe compensação para o explorado:
Os mecanismos de compensação podem ser importantes na estabilização da simbiose. Em geral, as simbioses são caracterizadas por uma diminuição na deriva genética e na taxa de especiação (Cf. efeito "Red King" ), mas nem sempre é o caso na micorrização: quanto mais a associação é exploradora, mais ela é. Específica porque a explorado desenvolve resistência que deve ser contornada pelo explorador ( Teoria da rainha vermelha ). No caso presente, a pressão seletiva no hospedeiro teria levado ao aparecimento de lignina e favorecido o crescimento dos tecidos vasculares.
Finalmente, a lógica e muitas pistas sugerem uma origem parasitária de micorrizas:
Existem dois tipos principais de micorrizas, definidas pelas relações fisioanatômicas entre os dois parceiros: endomicorrizas e ectomicorrizas. As micorrizas ericoides e aquelas associadas às orquídeas também são estudadas por seus serviços ecológicos , mas têm um interesse econômico mais limitado.
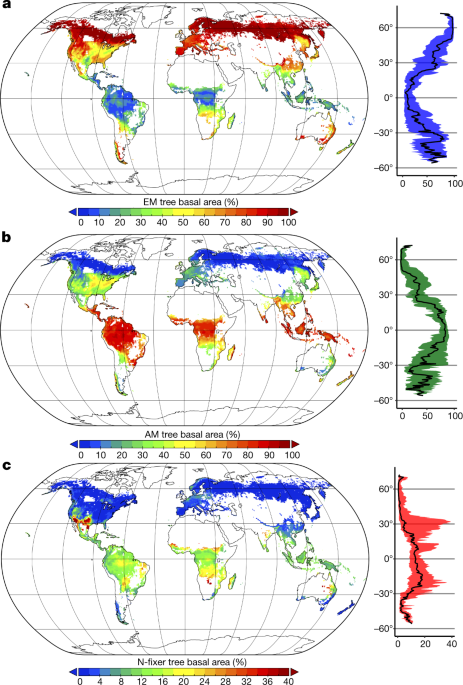
Um mapeamento global de simbioses florestais mostra o sucesso evolutivo das árvores ectomicorrízicas. Dos três tipos mais comuns de simbiose (endomicorrizas arbusculares abundantes em climas tropicais quentes e úmidos, ectomicorrizas em climas frios e bactérias fixadoras de nitrogênio em climas áridos e quentes), as ectomicorrizas representam apenas 2% das espécies de plantas, mas constituem 60% das todas as árvores do planeta.
Endomicorrizas (ou micorrizas internas) são a forma mais comum. São micorrizas que penetram nas raízes para se associarem melhor.
Existem vários tipos de endomicorriza:
Eles estão associados a plantas herbáceas e lenhosas. Essas endomicorrizas arbusculares, também chamadas de micorrizas vesiculares e arbusculares (ou micorrizas vesiculo-arbusculares), recebem seu nome de vesículas intercelulares (geralmente gotas de óleo, reserva de armazenamento para o fungo) e estruturas "intracelulares" que lembram uma pequena árvore. Se atravessarem bem a parede, entretanto, não penetram na membrana plasmática da célula vegetal, contentes em causar invaginação da membrana desta. Isso tem o efeito de aumentar a superfície de contato entre a hifa e a célula vegetal e, assim, facilitar a troca de metabólitos entre os dois parceiros.
As endomicorrizas arbusculares são formadas apenas por fungos da divisão Glomeromicetos que perderam a reprodução sexual. As hifas se estendem até o parênquima cortical da raiz, formando vesículas contendo reservas e estruturas ramificadas, os arbúsculos. Portanto, eles se reproduzem apenas assexuadamente. No entanto, as hifas de diferentes indivíduos podem se fundir, possibilitando a troca genética e uma forma de parassexualidade.
Eles também são geneticamente únicos, uma vez que seus esporos têm vários núcleos geneticamente diferentes.
Nessa simbiose, a densa e extensa rede de hifas de fungos micorrízicos ajuda a planta a explorar um volume maior de solo e ter acesso a locais inacessíveis para as raízes. O fungo permite que a planta melhore sua nutrição, fornecendo principalmente água, fósforo e nitrogênio . Aliás, a colonização das raízes por fungos micorrízicos permite protegê-las contra ataques de organismos patogênicos. Vários estudos mostraram que, sem associação com um fungo simbiótico, a planta crescerá mais lentamente (se crescer, como no caso do pinheiro preto em solo calcário) e terá muito mais probabilidade de ser vítima de uma infecção. Em contrapartida, o fungo se beneficia da fotossíntese da planta na forma de matéria orgânica rica em energia ( açúcares ) essencial à sua sobrevivência. A qualidade do húmus é assim melhorada, para benefício de outras espécies e para a manutenção ou constituição e melhoramento do solo.
Algumas plantas do Gimnosperma família, tais como Podocarpaceae , Araucariaceae e Phyllocladaceae , são conhecidos por ter inchaços raiz invadidas por fungos endomicorrízicos, chamados myconodules ou pseudonodules .
As ectomicorrizas (ou micorrizas externas) afetam apenas 5% das plantas vasculares, principalmente árvores de florestas temperadas e boreais (como Fagaceae , Pinacées ou Betulaceae ) e fungos da divisão de Ascomicetes , Basidiomicetos ou Zigomicetes . Essas micorrizas não penetram nas paredes celulares dentro das células da planta, mas simplesmente circundam as raízes, formando um manto de micélio e uma rede entre as paredes celulares da raiz.
O fungo associa-se primeiro a raízes finas com crescimento determinado, desprovido de pêlos radiculares. Em seguida, envolve a raiz de um manto de hifas , a manga micorrízica. Outras hifas crescem entre as células na parte externa do parênquima cortical, formando a interface simbiótica ou " rede de Hartig ". A simbiose modifica a fisionomia da raiz micorrízica: ela incha, pára de crescer e pode se ramificar abundantemente. O capuz e o meristema apical são então reduzidos.
Centenas de fungos diferentes podem estar associados à mesma espécie. A faia , por exemplo, tem um "recorde" com mais de 200 associações mychoriziennes.
O crescimento do micélio fúngico em direção às raízes da planta hospedeira é desencadeado pela percepção de um sinal emitido continuamente pelas raízes na forma de moléculas específicas da planta, como as estrigolactonas e, principalmente, os flavonóides .
Os sinais emitidos pelo fungo que permitem seu reconhecimento pela planta são mal compreendidos. Por analogia com os fatores Nod, esses fatores são chamados de "fatores Myc". Moléculas semelhantes ao fator Nod produzidas por fungos arbusculares e tendo um efeito na planta hospedeira foram recentemente demonstradas, mas seu papel na sinalização simbiótica ainda precisa ser determinado.
A via de sinalização ativada pelo fator myc tem etapas em comum com a via Nod presente em leguminosas noduladas por rizóbio e em plantas actinorrízicas . Os receptores dos fatores Myc não são conhecidos, mas as proteínas que intervêm posteriormente, como SYMRK, CASTOR, POLLUX, NUP, CYCLOPS, são necessárias para as simbioses micorrízicas rizobiais e arbusculares. Como na simbiose rizobiana, o contato com um fungo arbuscular induz oscilações de cálcio. Como a endomicorriza arbuscular apareceu antes da endossimbiose fixadora de nitrogênio, os pesquisadores levantaram a hipótese de que a via de transdução do sinal micorrízico foi recrutada por bactérias fixadoras de nitrogênio.
Uma procissão micorrízica ideal atuaria como um "fertilizante" sem seus inconvenientes e gratuitamente, sem pesticidas , fungicidas e aditivos . Após os testes iniciais na Austrália na década de 1920, as empresas desenvolveram cepas de inoculação adequadas para várias espécies comerciais (inóculos de uma ou várias camadas), mas também precisariam ser adaptadas ao contexto do local. Vários pesquisadores acreditam que as cepas de fungos simbiontes geneticamente mais adaptados ao campo são aquelas que são indígenas. Assim, empresas privadas em todo o mundo desenvolveram a produção e venda de inoculantes micorrízicos destinados ao uso agrícola ( biofertilizante , bioestimulante ), mas também ecológico ( revegetação de solos erodidos, fitorremediação ).
Na agricultura, o uso de fungos micorrízicos evitaria muitos problemas de desnaturação de solos orgânicos. Com efeito, a adição de pesticidas e, em particular, de fungicidas pode ter efeitos nocivos na qualidade do solo. Os fungos têm papéis importantes na manutenção do solo fértil. A adição de compostos químicos com alto teor de fósforo induziria ao esgotamento da quantidade e da qualidade dos fungos endomicorrízicos, reduzindo o aporte de nutrientes às plantas cultivadas. Além disso, o excesso de fósforo tem um impacto negativo nas vias navegáveis (eutrofização). Portanto, é essencial reduzir a quantidade de produtos químicos nocivos e, em vez disso, optar pela inoculação de fungos naturais que teriam as mesmas vantagens de um crescimento rápido, sem as desvantagens para o meio ambiente.
Um estudo em jujuba demonstrou que a micorrização controlada foi benéfica para o seu crescimento e nutrição com fosfato. Na verdade, a árvore frutífera absorvia o fósforo natural adicionado em solo pobre com muito mais facilidade. Ele até dependia disso para um crescimento normal. Portanto, não basta, na agricultura, simplesmente adicionar fósforo em grandes quantidades para que a planta cresça mais rápido, é também uma questão de levar em conta a rede micorrízica. Em primeiro lugar, deve ser capaz de absorver com eficácia. Uma overdose de nutrientes não mudaria nada e só causaria a lixiviação do fósforo e a poluição das águas superficiais.
A ação das micorrizas não para com uma melhor nutrição: elas também permitem que a planta se proteja melhor contra fatores bióticos e abióticos prejudiciais. É claro que uma nutrição melhor implica uma melhor saúde da planta, que pode então tolerar melhor os estresses ambientais, em particular os patógenos. As micorrizas podem liberar antibióticos na rizosfera que têm o efeito de eliminar todos os microorganismos, patogênicos ou não, que podem desviar parte do comércio em seu benefício. Além disso, a simbiose micorrízica também induz o estabelecimento de mecanismos de defesa na própria planta (imunidade e estimulação imunológica). É uma proteção indireta localizada a nível celular. Em plantas ectomicorrízicas , as células de rizoderma sintetizam taninos armazenados em vacúolos que fornecem proteção contra novos ataques de micróbios e outros patógenos. A planta também produz mais lignina nas paredes celulares do endoderma e nos tecidos vasculares. Também pode induzir ou suprimir várias defesas ligadas a fitoalexinas , peroxidases , quitinases e várias outras. Em suma, o fungo provoca mudanças consideráveis nos mecanismos de defesa da planta contra diversos parasitas, e isso antes das infecções.
Às vezes, a associação com um fungo causa uma remodelação do sistema radicular. Por exemplo, em morangos, a micorrização causa maior proteção contra o apodrecimento da raiz causado por Phytophthora fragariae . A intensa ramificação das raízes induzida pelo fungo micorrízico é acompanhada por exsudatos radiculares, e isso teria o efeito de modificar a microflora e as interações diretas com os parasitas.
Em relação aos fungos parasitas, uma planta associada a uma micorriza também está mais protegida. As hifas do fungo simbiótico colonizam as raízes da planta e "bloqueiam" os locais de acesso do fungo parasita. Eles também competem com os nutrientes disponíveis. Uma micorriza também recebe benefícios ao se ligar a uma planta: ela recebe compostos de carbono em troca de minerais. Visto que o carbono é altamente cobiçado, uma micorriza tem uma vantagem maior do que um fungo parasita livre no solo. Além disso, as micorrizas podem representar até 80% da massa microbiana do solo. Portanto, eles influenciam muito as propriedades físicas e químicas do ambiente e podem, portanto, controlar várias interações microbianas no solo. Às vezes está de acordo com a planta micorrízica, proporcionando proteção e crescimento, mas às vezes os efeitos são bastante negativos.
As micorrizas podem, portanto, ajudar a proteger a planta de estresses abióticos (por exemplo, seca) e bióticos de várias maneiras: melhor nutrição e saúde, transformação morfológica das raízes, indução ou supressão de mecanismos de defesa e agindo sobre o próprio parasita, seja por competição por recursos e locais de infecção, ou por modificação da microflora e um aumento no nível de matéria orgânica.
Deve-se notar, no entanto, que essas interações são estudadas em estufas e em ambientes controlados e que, portanto, não refletem totalmente toda a complexidade de um ambiente natural que varia no tempo e na composição.
Com todos os benefícios declarados das micorrizas, é justo dizer que seu uso adequado evitaria muitas preocupações ambientais, pois atuariam como fertilizantes e agentes protetores. O uso de fertilizantes químicos, fungicidas e pesticidas não seria tão necessário. Na verdade, os fertilizantes diminuem a taxa de micorrização, o que tem o efeito de aumentar a dependência das plantas desses insumos. Da mesma forma, o uso de agrotóxicos diminui o poder inoculante do solo, resultando na diminuição do efeito protetor das micorrizas e no aumento da demanda das plantas por agrotóxicos. A adição de tais entradas configura uma forma de círculo vicioso .
A atividade silvicultural e agrícola pode interromper ou modificar (negativa ou positivamente) a microflora fúngica e sua capacidade de micorrizar;
Mudanças bioquímicas e biológicas, bem como alterações microclimáticas induzidas por grandes cortes rasos, (ou mesmo por colheitas no caso da agricultura) podem ter impactos subestimados, ligados a impactos diretos no solo, na água, mas também à perda de fungos inóculo (mesmo que as raízes permaneçam no solo, a " madeira grande " e a " grande madeira morta " tornam-se raras ou ausentes, e muitas vezes o corte é recolhido).
O termo micorriza (do grego myco , “fungo” e riza , “raiz”) foi introduzido em 1885 pelo botânico Albert Bernhard Frank . Contratado pelo rei prussiano Guilherme I st para desenvolver métodos de produção de trufas estudando como elas se espalham, ABFrank observa que estes fungos associado com as raízes de árvores e propõe que esta associação é uma relação simbiótica e não parasitária . A teoria da ecologia evolutiva de Frank, contradizendo a crença de que trufas e outros fungos causam doenças e podridões em plantas, é fortemente contestada por seus colegas. Essa teoria retorna ao centro das atenções com o trabalho de Jack Harley considerado o "papa das micorrizas" por seus estudos desde 1950, Barbara Mosse em 1962, JM Phillips e DS Hayman em 1970, M. Gardes e TD Bruns em 1993.
A pesquisa está se desenvolvendo rapidamente no início do XXI th século com novos meios de biologia molecular e genética , a ponto de dar o nome a uma disciplina científica , o mycorhizologie , estudado por mycorhizologues . De 2008 a 2016, mais de 10.000 novas contribuições científicas trataram de micorrizas.
{kind=link}